 I use a comparative approach to investigate the evolution and development (evo-devo) of shoot architecture. Typically, evo-devo studies seek to understand the molecular and developmental basis of morphological change. These studies have focused primarily on macroevolutionary problems by comparing fixed developmental patterns across broad phylogenetic distances, often between model organisms. Broad-scale comparative studies have identified key gene networks that control the morphological differences between major clades. Presumably, changes in these developmental pathways also are responsible for population level variation, but whether they actually are - and how these changes manifest - is less clear. In light of this, a recent NSF workshop on Frontiers in Evolutionary Biology recognized the need to integrate development and microevolution. My approach to this problem is to re-evaluate the same gene networks that produce morphological differences among clades, but at the population level where developmental variation arises and is subject to microevolutionary processes.
I use a comparative approach to investigate the evolution and development (evo-devo) of shoot architecture. Typically, evo-devo studies seek to understand the molecular and developmental basis of morphological change. These studies have focused primarily on macroevolutionary problems by comparing fixed developmental patterns across broad phylogenetic distances, often between model organisms. Broad-scale comparative studies have identified key gene networks that control the morphological differences between major clades. Presumably, changes in these developmental pathways also are responsible for population level variation, but whether they actually are - and how these changes manifest - is less clear. In light of this, a recent NSF workshop on Frontiers in Evolutionary Biology recognized the need to integrate development and microevolution. My approach to this problem is to re-evaluate the same gene networks that produce morphological differences among clades, but at the population level where developmental variation arises and is subject to microevolutionary processes.
I address this problem by examining the molecular and developmental basis for intraspecific variation of shoot architecture in two populations of Mimulus guttatus (monkeyflower). Branching, the production of new axes of growth, is a developmental process that has profound consequences for plant morphological and life history evolution. In Oregon, M. guttatus plants from the coastal dunes flower late, are highly branched, and often overwinter (i.e. exhibit a perennial life-cycle). In contrast, individuals from Iron Mountain, an alpine habitat in the Cascades, flower early, have fewer branches, and exhibit an annual life cycle. Despite these clearly documented differences, the developmental and evolutionary dynamics behind this adaptive divergence in life history are unknown. My dissertation will determine whether gene networks responsible for generating differences in shoot architecture in broadly divergent taxa are also responsible for population level variation in M. guttatus.

Mimulus guttatus is found in the open slopes and spring seeps along the flanks of Iron Mountain in the Cascade range of Oregon. This is the same population that the genome sequence for M. guttatus comes from.
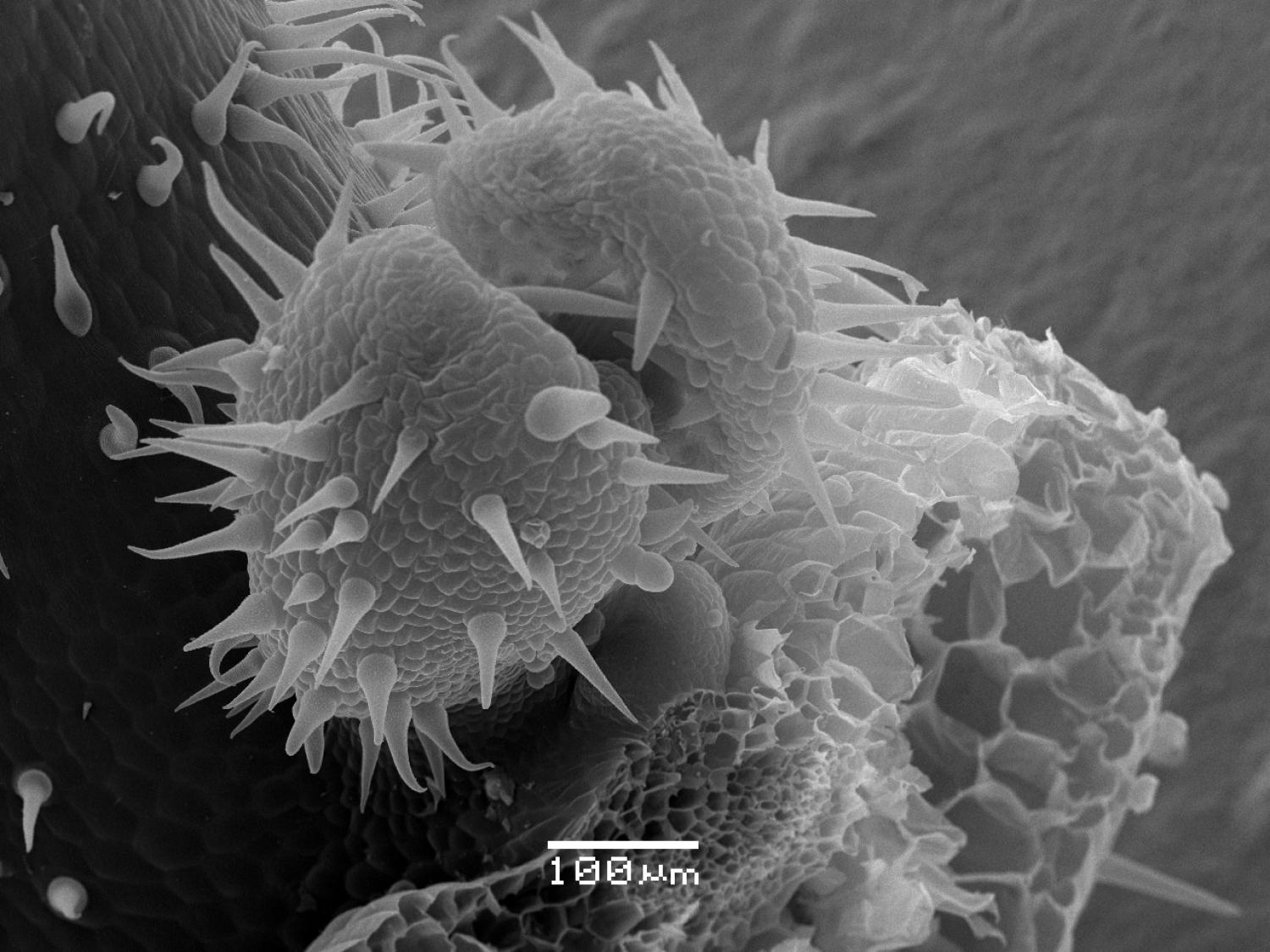
From left to right, the main stem, axillary bud, secondary axillary meristem, and subtending leaf of at the first node of a young M. guttatusplant from Iron Mountain. Iron Mountain plants tend to have very few branches, nevertheless they initiate multiple axillary meristems per node. In their natural environment, these buds probably wouldn’t have the chance to develop into branches.

Iron Mountain M. guttatus plants flower young in the field too.

An experiment in progress. M. guttatus plants are highly phenotypically plastic. Therefore, I grow them in growth chambers. This minimizes variation between populations that is due to environmental noise and allows me to tease apart the genetic differences between the populations.





